فلورتین با فعال سازی Nrf2 به واسطه اتوفاژی در ماکروفاژها التهاب عصبی را سرکوب می کند.
Mar 13, 2022
مخاطب:tina.xiang@wecistanche.com
خلاصه
زمینهماکروفاژها نقش دوگانه ای در اختلالات عصبی التهابی مانند ام اس دارند. آنها در شروع و پیشرفت ضایعه نقش دارند، اما می توانند باعث بهبودی نیز شوندالتهابو ترمیم بافت آسیب دیده در این مطالعه به بررسی اینکه آیا و چگونه می پردازیمفلورتینفلاونوئیدی که به وفور در سیب و توت فرنگی وجود دارد، فنوتیپ التهابی ماکروفاژها را کاهش داده و سرکوب می کند.التهاب عصبی.
مواد و روش هاتغییرات رونویسی در ماکروفاژهای مشتق از مغز استخوان موش پس از مواجهه با فلورتین با توالی RNA حجیم ارزیابی شد. مسیرهای زمینهای مرتبط با التهاب، پاسخ استرس اکسیداتیو و اتوفاژی با روشهای کمی PCR، فلورسنت و جذب، موشهای ناک اوت مرتبط با فاکتور 2 (Nrf2)، وسترن بلات و ایمونوفلورسانس تأیید شدند. مدل تجربی آنسفالومیلیت خودایمنی (EAE) برای مطالعه تأثیر فلورتین بر التهاب عصبی در داخل بدن و تأیید مکانیسمهای زمینهای مورد استفاده قرار گرفت.
نتایجما نشان میدهیم که فلورتین فنوتیپ التهابی ماکروفاژها را کاهش میدهد و به طور قابل توجهی التهاب عصبی را در EAE سرکوب میکند. فلورتین اثر خود را با فعال کردن مسیر سیگنالینگ Nr2 واسطه می کند. فعالسازی Nrf2 به فعالسازی وابسته به پروتئین کیناز 5'AMP (AMPK) در اتوفاژی و متعاقب آن تخریب پروتئین 1 (Keap1) مرتبط با ECH مانند کلچ نسبت داده شد.
نتیجه گیری: این مطالعه چشم اندازهای آینده را برای فلورتین به عنوان یک استراتژی درمانی برای اختلالات عصبی التهابی مانند ام اس باز می کند.
ثبت نام آزمایشی: قابل اجرا نیست
کلید واژه ها: فلورتین، ماکروفاژها، التهاب عصبی، مولتیپل اسکلروزیس، خودایمنی

برای کسب اطلاعات بیشتر اینجا را کلیک کنید
زمینه
ضایعات مولتیپل اسکلروز فعال (MS) با وجود ضایعات متعدد مشخص می شودماکروفاژها[1-5]. مطالعات اولیه نشان داد که ماکروفاژها یک فنوتیپ پیش التهابی را در ضایعات MS اتخاذ می کنند و در نتیجه باعث افزایشالتهاب عصبی، دمیلیناسیون و تخریب عصبی. توابع موثر تحریک کننده بیماری شامل ارائه آنتی ژن های مشتق از سیستم عصبی مرکزی (CNS) به سلول های T خودواکنشی و تولید واسطه های التهابی مانند سیتوکین های پیش التهابی، گونه های اکسیژن فعال (ROS) و اکسید نیتریک (NO) است. 6،7]. اخیراً مشخص شد که ماکروفاژها همچنین عملکردهای مفیدی در ضایعات MS دارند زیرا می توانند فنوتیپ خود را به فنوتیپ تغییر شکل دهند که معمولاً با سرکوب سیستم ایمنی و ترمیم CNS مرتبط است. این فنوتیپ محافظ با کاهش تولید واسطه های پیش التهابی، تولید واسطه های ضد التهابی و فاکتورهای رشد، و فعال شدن مسیرهای ضد اکسیداتیو مانند مسیر فاکتور هسته ای اریتروئید 2-مربوط به فاکتور 2 (Nrf2) مشخص می شود. [8-14]. از آنجایی که وضعیت التهابی ماکروفاژها با پیشرفت MS و ایجاد ضایعات فعال مزمن مرتبط است، هدایت ماکروفاژها به یک فنوتیپ مفید به عنوان یک استراتژی امیدوارکننده برای محدود کردن پیشرفت MS در نظر گرفته می شود [15].
اجزای رژیم غذایی باعث عملکرد ماکروفاژها و التهاب عصبی می شوند [16]. به طور خاص، خانواده فلاونوئیدها به طور فزاینده ای دارای ترکیبات امیدوارکننده ای هستند که بر مسیرهای بیماری زا تأثیر می گذارند و فنوتیپ سلول های ایمنی مانند ماکروفاژها را تعدیل می کنند [17،18]. فلاونوئیدها یکی از بزرگترین خانواده های گیاهی را تشکیل می دهند که حاوی بیش از 8000 ترکیب فنلی با فعالیت زیستی متنوع است. چندین عضو از خانواده فلاونوئیدها اثرات ضد التهابی و ضد اکسیداتیو را روی ماکروفاژها نشان میدهند [19-21]. فلاونوئید فلورتین یکی از اعضای دی هیدروکالکون ها است و در میوه های معمولی مانند سیب و توت فرنگی وجود دارد. فلورتین دارای ویژگیهای تعدیلکننده ایمنی است و به دلیل ویژگی آنتیاکسیدانی که دارد [{8}}] به طور گسترده برای مراقبت از پوست استفاده میشود. علاوه بر این، فلورتین یک مهارکننده ناقل گلوکز (GLUT) است، ویژگی که فنوتیپ ماکروفاژها را تحت تاثیر قرار می دهد زیرا فعال شدن ماکروفاژها توسط GLUT تحریک می شود [25، 26]. به طور کلی، این ویژگی ها فلورتین را به ترکیبی امیدوارکننده برای تعدیل فنوتیپ ماکروفاژها و التهاب عصبی تبدیل می کند. در این مطالعه، ما نشان میدهیم که فلورتین ماکروفاژها را به سمت فنوتیپ کمتر التهابی سوق میدهد و التهاب عصبی را در مدل آنسفالومیلیت خودایمنی تجربی (EAE) کاهش میدهد. توالی یابی RNA و آزمایش های عملکردی مسیر Nrf2 را به عنوان یک هاب و محرک این فنوتیپ ماکروفاژ محافظ القا شده توسط فلورتین شناسایی کرد. فعالسازی دستگاه اتوفاژی وابسته به AMPK و متعاقب آن SQSTMl/p{14} تخریب Keapl، آداپتوری که تخریب پروتئازومی Nrf2 را تسهیل میکند، با واسطه (که از این به بعد p62 نامیده میشود)، زمینه ساز فعالسازی Nrf2 در ماکروفاژها است. این یافته ها نشان می دهد که فلورتین پتانسیل استفاده در استراتژی های درمانی اختلالات التهابی عصبی را دارد.
مواد و روش ها
آنتی بادی ها و معرف های شیمیایی
فلورتین(سیگما آلدریچ) در 5{13}} میلی مولار KOH به محلول ذخیره 15 میلی مولار حل شد و در درجه{2}} نگهداری شد. رقت های بیشتر در محیط RPMI1640 (Gibco) ساخته شد. برای درمان in vivo، فلورتین در 1 N NaOH حل شد، سپس pH با 1 N HCL به 7.2 تنظیم شد و محلول بیشتر در آب فیزیولوژیکی رقیق شد تا غلظت 50 میلی گرم بر کیلوگرم به دست آید. BML{9}}(1 میکرومولار، بیوتکنولوژی سانتا کروز) 1 ساعت قبل از درمان با فلورتین برای مهار فعالسازی AMPK اضافه شد. بافیلومایسین A1 (BAF، 0.1 میکرومولار، InvivoGen) 2 ساعت قبل از جمع آوری اضافه شد تا از همجوشی اتوفاگوزوم ها و لیزوزوم ها جلوگیری کند. لیپوپلی ساکارید (LPS، 100 نانوگرم در میلی لیتر، Sigma-Aldrich) برای تحریک سلول ها برای فنوتیپ التهابی استفاده شد. فوربول 12-میریستات 13-استات (PMA، 100 نانوگرم در میلی لیتر، سیگما آلدریچ) برای القای تولید ROS استفاده شد. آنتیبادیهای زیر برای وسترن بلات استفاده شد: آنتی{22}}اکتین موش (1:10،000;sc-47778، بیوتکنولوژی سانتا کروز)، ضد GAPDH موش (1:{29) }};AB_2537659، Invitrogen)، ضد AMPK خرگوش (1:1000; 5831S، فناوری سیگنالینگ سلولی)، AMPK ضد فسفریله خرگوش (1:1000؛ 2535S، فناوری سیگنالینگ سلولی)، خرگوش ضد LC3 (1:1000؛ L7543، سیگما-آلدریچ)، خرگوش anti-p62 (1:1000؛ 23214، فناوری سیگنالینگ سلولی). آنتی بادی های زیر برای ایمونوفلورسانس استفاده شد: ضد CD3 موش (1:150؛ MCA500G، Bio-Rad)، موش anti-F4/80 (1:100؛ MCA497G، Bio-Rad)، خرگوش anti-LC3 (1:1000) ;L7543, Sigma-Aldrich), rabbit anti-p62 (1:500;23214, Cell Signaling Technology), rabbit anti-Keap1(1:500;60027-1-Ig,Proteintech Europa), rabbit anti-TMEM119(1) :100, ab209064, Abcam). آنتی بادی های ثانویه مناسب از Invitrogen خریداری شد.
موش
موش های نوع وحشی (WT) C57BL/6JOlaHsd از Envigo خریداری شدند. حیوانات با رژیم غذایی منظم تغذیه شدند و در مرکز حیوانات موسسه تحقیقات زیست پزشکی دانشگاه هاسلت نگهداری شدند. تمام آزمایشها بر اساس دستورالعملهای سازمانی انجام شد و توسط کمیته اخلاقی آزمایشهای حیوانی دانشگاه هاسلت تأیید شد.
کشت سلولی
مشتق از مغز استخوانماکروفاژها(BMDMs) از موشهای WT و Nrf2 Knockout (KO) C57BL/6JOlaHsd، که از Envigo خریداری و توسط RIEN BRC بر اساس MTA به پروفسور S. Kerdine-Römer ارائه شد، جدا شدند [27،28]. BMDM ها همانطور که قبلا توضیح داده شد به دست آمدند [29]. به طور خلاصه، سلولهای مغز استخوان درشت نی و استخوان ران از موشهای 12- هفتهای WT و Nrf2 KO C57BL/6JOlaHsd در پلیتهای پتری 10- سانتیمتری با غلظت 10×106 سلول/ کشت داده شدند. پلیت، در محیط RPMI1640 همراه با 10 درصد سرم جنین گوساله (FCS, Gibco)، 50U/ml پنی سیلین (Invitro-gen)، 50 U/ml استرپتومایسین (Invitrogen) و 15 درصد L929-محیط آماده شده (LCM) ). پس از تمایز، BMDM ها در دمای 37 درجه با 10 میلی مولار EDTA در PBS (Gibco) جدا شدند و (0.5×10 درجه سلول در میلی لیتر) در RPM1640 همراه با 10 درصد FCS، 50U/ml پنی سیلین، 50U/ml استرپتومایسین و 5 درصد LCM کشت داده شدند. 37 درجه سانتی گراد، 5 درصد CO2). کشت میکروگلیا از مغز نوزادان P{36}} C57BL/6/OlaHsd پس از تولد جدا شد. پس از برداشتن ساقه مغز، شبکه مشیمیه و مننژها، مغزها به صورت مکانیکی جدا شده و به مدت 15 دقیقه با تریپسین 1× (گیبکو) در دمای 37 درجه هضم شدند. پس از آن، سوسپانسیون سلولی در DMEM یک محیط با گلوکز بالا (سیگما) همراه با 30 درصد LCM، 10 درصد FCS، 50U/ml پنی سیلین و 50U/ml استرپتومایسین در فلاسک های کشت T75 کشت شد. دو تا سه روز بعد، تغییر کامل محیط انجام شد. کشت گلیال مخلوط (230rpm، 3h، 37 درجه) پس از 6-7 روز تکان داده شد تا کشت میکروگلیا خالص بدست آید.
توالی یابی RNA
سلول ها با فلورتین (50 میکرومولار) به مدت 20 ساعت و تحریک LPS (100 نانوگرم در میلی لیتر) به مدت 6 ساعت پیش تیمار شدند. لیز سلولی با استفاده از معرف Qiazol Lysis (Qiagen) انجام شد. RNA با استفاده از کیت کوچک RNeasy (Qiagen) از سلول ها استخراج شد. سپس نمونه ها توسط Genomics Core Leuven (بلژیک) پردازش شدند. آماده سازی کتابخانه با کیت QuantSeq Lexogen برای تولید کتابخانه های سازگار با Illumina انجام شد. کتابخانه ها بر اساس سیستم توالی یابی Illumina HiSeq4000 توالی یابی شدند. هم ترازی Splice-aware با STAR v2.6.1b [30] انجام شد. کمیت خواندن در هر ژن با HT-seq Count v2.7.14 انجام شد. تجزیه و تحلیل بیان دیفرانسیل مبتنی بر شمارش با بسته زیست رسانا DESeq2 مبتنی بر R (بنیاد R برای محاسبات آماری، وین، اتریش) انجام شد. لیستی از ژن های متفاوت بیان شده در ap انتخاب شد<0.05 and="" used="" as="" an="" input="" for="" the="" core="" analysis="" by="" qiagen's="" ingenuity="" pathway="" analysis="" (ipa).="" all="" rna="" sequencing="" (rna-seq)="" data="" discussed="" in="" this="" publication="" have="" been="" deposited="" in="" ncbi's="" gene="" expression="" omnibus(edgar="" et="" al,="" 2002)="" and="" are="" accessible="" through="" geo="" series="" accession="" number="">
رونویسی معکوس کمی PCR
سلول ها با فلورتین (5{8}} میکرومولار) به مدت 20 ساعت و LPS (100 نانوگرم در میلی لیتر) به مدت 6 ساعت پیش تیمار شدند. لیز با استفاده از معرف Qiazol Lysis (Qiagen) انجام شد. RNA با استفاده از کیت کوچک RNeasy (Qiagen) استخراج شد. غلظت و کیفیت RNA با اسپکتروفتومتر قطره ای نانو (Isogen Life Science) تعیین شد. سنتز cDNA با استفاده از Quanta qScript cDNA SuperMix (Quanta Biosciences) طبق دستورالعمل سازنده انجام شد. qPCR بر روی یک سیستم Real-Time PCR StepOne-Plus (Applied Biosystems) با استفاده از مخلوط سبز رنگ SYBR حاوی 1× SYBR سبز (Applied Biosystems)، پرایمرهای 0.3 میکرومولار (تکنولوژی های DNA یکپارچه)، 12.5 نانوگرم cDNA-free آب و هسته انجام شد. برای تعیین کمیت بیان ژن از روش مقایسه ای Ct استفاده شد.داده ها به پایدارترین ژن های مرجع سیکلین A و هیپوگزانتین فسفریبوزیل ترانسفراز 1 نرمال سازی شدند. توالی های پرایمر در صورت درخواست در دسترس هستند.
تعیین گونه های اکسیژن فعال
سلول ها با فلورتین (50 میکرومولار) به مدت 2 ساعت پیش تیمار شدند. پس از آن، سلول ها با PMA (15 دقیقه، 100 نانوگرم در میلی لیتر) تحریک شدند و تولید ROS با استفاده از پروب فلورسنت 2',7'-dichlorodihydrofluorescein diacetate در 10 میکرومولار در PBS به مدت 30 دقیقه اندازه گیری شد. فلورسانس با استفاده از میکروپلیت خوان فلورسانس FLUOstar optima (BMG Labtech، Ortenberg، آلمان) اندازه گیری شد (تحریک: 495 نانومتر، انتشار: 529 نانومتر).
اندازه گیری اکسید نیتریک
سلول ها با فلورتین (50 میکرومولار) به مدت 2 ساعت پیش تیمار شدند. پس از آن، سلول ها با LPS (24 ساعت، 100 نانوگرم در میلی لیتر) تحریک شدند. NO به طور غیر مستقیم با استفاده از کیت اندازه گیری نیتریت معرف Griess (Abcam) پایش شد. به طور خلاصه، نیترات با سولفانیل آمید و N-(1-نفتیل)اتیلن-دیامین دی هیدروکلراید واکنش می دهد تا رنگ آزو صورتی تولید کند. سپس جذب این مشتق آزو با استفاده از دستگاه میکروپلیت خوان (iMark، Bio-Rad) در طول موج 540 نانومتر اندازه گیری شد.
وسترن بلات
سلول ها با فلورتین (5{{1{18}}}} میکرومولار) و LPS (100 نانوگرم در میلی لیتر) به مدت 1 ساعت یا 24 ساعت برای تعیین میزان فعال سازی AMPK یا سطوح پروتئین p62، LC3 و Keapl تحت درمان قرار گرفتند. سلول ها با استفاده از بافر RIPA (150 میلی مولار NaCl، 1 درصد تریتون X{9}}، 0.5 درصد دی اکسی کولات سدیم، 1 درصد SDS، 50 میلی مولار تریس) لیز شدند و از طریق الکتروفورز ژل پلی آکریل آمید دودسیل سولفات سدیم جدا شدند. ژلها به غشای PVDF (VWR) منتقل شدند و بلاتها به مدت 1 ساعت در آلبومین سرم گاوی 5 درصد در سالین بافر Tris حاوی 0.1 درصد Tween{20}} (TBS-T) مسدود شدند. غشاها با آنتیبادیهای اولیه یک شبه در دمای 4 اینچ بررسی شدند، با TBS-T شسته شدند و با آنتیبادی ثانویه نشاندار شده با پراکسیداز ترب کوهی به مدت 1 ساعت در دمای اتاق (RT) انکوبه شدند. ترمو فیشر با استفاده از Amersham Imager 680 (GE Healthcare Life Sciences) چگالی باندها با استفاده از ImageJ تعیین شد.
ایمونوفلورسانس
انجمادهای نخاعی در هوا خشک و در استون سرد به مدت 10 دقیقه در درجه {3}} ثابت شدند. BMDM های موش بر روی لام های پوشش شیشه ای کشت و در متانول سرد به مدت 10 دقیقه در درجه {6}} ثابت شدند. برش ها و BMDM ها با استفاده از بلوک پروتئین داکو (Agi-lent) به مدت 30 دقیقه مسدود شدند. پس از آن، آنها یک شبه در دمای 4 سانتیگراد با آنتیبادیهای اولیه انکوبه شدند، شسته شدند و با آنتیبادیهای ثانویه مناسب به مدت 1 ساعت در RT انکوبه شدند. نرمافزار Elements BR 3.10 (Nikon) تصاویر BMDM رنگآمیزی شده برای p62، LC3 و Keapl با استفاده از میکروسکوپ کانفوکال Zeiss LSM 880 گرفته شد و با تصحیح Airyscan (63× عینی).P{19}}و LC{20}} نقطه های مثبت با استفاده از آنالیز نقطه ای نیمه خودکار imageJ تعیین شد.به طور خلاصه، پس از اینکه تصویر باینری شد و سلول ها با دست انتخاب شدند، نقاط پونکتا در هر سلول آنالیز شدند. همون سازی P62 و Keapl با استفاده از خط لوله colocalization در نرم افزار CellProfiler انجام شد [31]. ]. تصاویر نشان داده شده در شکل ها به صورت دیجیتالی بهبود یافته اند.
Experimental autoimmune encephalomyelitis model Eleven-week-old C57BL/6JOlaHsd mice were immunized subcutaneously with 200 ng of myelin oligodendrocyte glycoprotein peptide(MOG35-55)emulsified in 100 ul complete Freund's supplemented with 4 mg/ml of Mycobacterium tuberculosis(EK2110, Hooke Laboratories). Im-mediately after MOG immunization and after 24 h, mice were intraperitoneally injected with 50 ng pertussis toxin (EK2110 kit, Hooke Laboratories) to induce EAE.EAE animals were treated daily with phloretin or vehicle(50 mg/kg, intraperitoneal (ip)) after 6 days of immunization (prophylactic setup)or after disease onset(clinical score>1، تنظیم درمانی). موشها طبق راهنمای امتیازدهی EAE موشهای سازنده روزانه وزن و نمرهگذاری شدند تا علائم عصبی بیماری را نشان دهند:0:بدون علائم بالینی،0.5: نوک دم شل است، 1: دم شل است. 1.5: لنگی دم و بازداری پای عقب 2: لنگی دم و ضعف پاهای عقبی 2.5: لنگی دم و کشیدن پاهای عقبی 3: لنگی دم و فلج کامل پاهای عقبی 3.5: لنگی دم و فلج کامل عقبی پاها و موش وقتی روی پهلو قرار می گیرند قادر به درست کردن خود نیستند، 4: فلج دیافراگم، 5: مرگ توسط EAE.
تحلیل آماری
برای تجزیه و تحلیل آماری دادهها از GraphPad Prism استفاده شد که به صورت میانگین ± D'Agostino نشان داده میشوند و برای آزمون توزیع نرمال از آزمون نرمال بودن omnibus Pear-son استفاده شد. برای داده های توزیع شده نرمال از آزمون t-test دانشجوی بدون جفت دو طرفه (در صورت لزوم با تصحیح ولش) استفاده شد. برای داده هایی که آزمون نرمال بودن را قبول نکردند، از تحلیل من ویتنی استفاده شد<0.05 were="" considered="" to="" demonstrate="" significant=""><><0.01,><0.001 and="" *p=""><>

نتایج
تغییرات رونویسی مرتبط با درمان فلورتین ماکروفاژها
برای ایجاد اثرات ضد التهابی بالقوه و شناسایی مکانیسمهای اساسی درمان فلورتین روی ماکروفاژها، توالییابی RNA حجیم را انجام دادیم (شکل تکمیلی 1). تجزیه و تحلیل مسیر ماکروفاژهای فعال تیمار شده با فلورتین نشان داد که ژن های بیان شده متفاوت در مسیرهای متعارف مربوط بهالتهاب، مانند iNOS(z-score:-2.449)، گیرنده شبه عوارض (z-score:-2.236)، سیگنالینگ اینترفرون (z-score:- 2)، و مسیر پاسخ فاز حاد (z-score:{10}}.633)(Fig.1A, B). مشابه تجزیه و تحلیل مسیر متعارف، تجزیه و تحلیل بالادستی دادههای RNA-seq پیشبینی کرد که فلورتین فعالسازی تنظیمکنندههای کلیدی رونویسی پیش التهابی مانند IRF7 (z-score:2.229)، IRF1 (z-score:-2) را کاهش میدهد. 025)و STAT1(z-score:-2.022)(شکل 1D). در کنار کاهش مسیرهای پیش التهابی ماکروفاژها، تجزیه و تحلیل RNA-seq ماکروفاژهای تیمار شده با فلورتین نشان داد که در میان مسیرهای دیگر، فلورتین به شدت مسیر Nrf2 را فعال می کند (z-score: 1.897)، که توسط دخیل در Nrf مرتبط می شود{35} ژن هایی مانند mafG و پروکسی (شکل 1A, C). حتی بیشتر، Nrf2 (NFE2L2) به عنوان فعالترین تنظیمکننده رونویسی بالادست (z-score: 2.801) و تنظیم سطوح ROS به عنوان یکی از تنظیمشدهترین عملکردهای بیولوژیکی در BMDMهای تحت درمان با فلورتین شناسایی شد (z-score: 2.008). )(شکل 1D، E). در مجموع، یافته ها نشان می دهد که فلورتین Nrf2 را فعال می کند و فنوتیپ التهابی ماکروفاژها را سرکوب می کند.
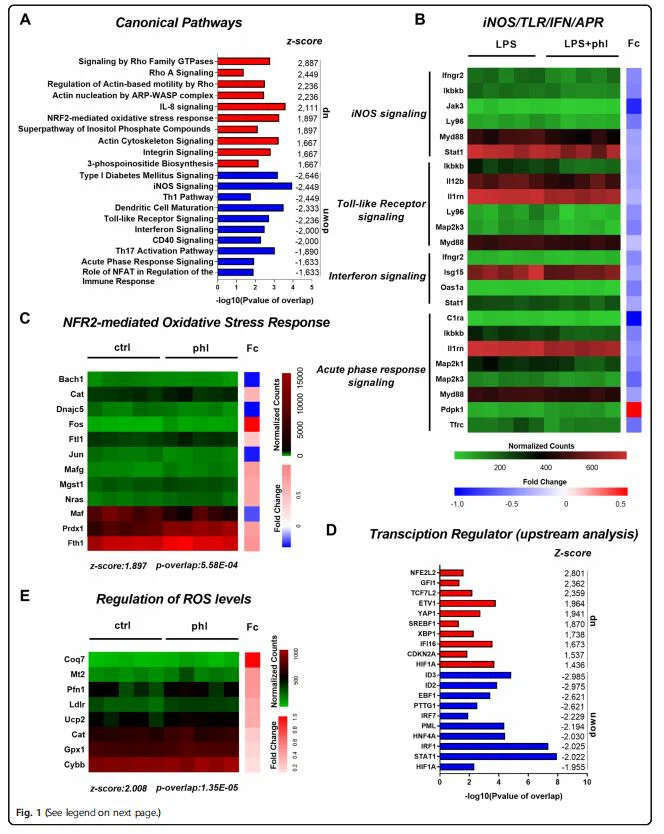
مسیر Nrf2 فنوتیپ ماکروفاژهای تیمار شده با فلورتین را کنترل می کند.
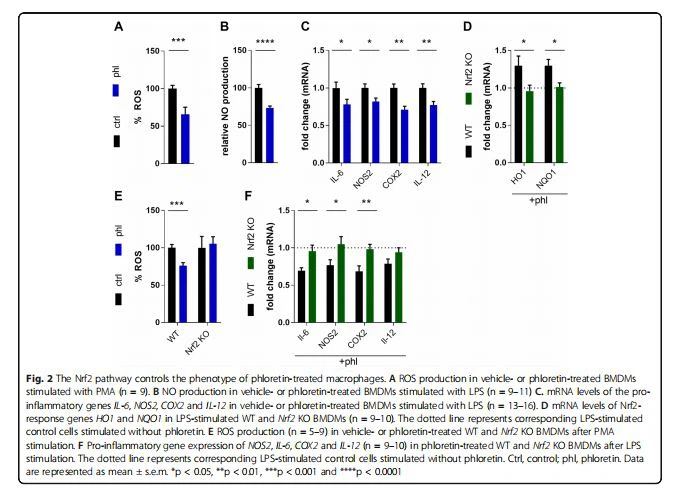
در مرحله بعد، ما اثر ضد التهابی فلورتین را تأیید کردیم و تعیین کردیم که آیا فنوتیپ التهابی ماکروفاژها را با تأثیر بر Nrf2 تعدیل می کند یا خیر. کاهش سطح ROS در WT BMDMهای تیمار شده با فلورتین تحریک شده با PMA مشاهده شد (شکل 2A). علاوه بر این، کاهش تولید NO و سطوح mRNA ژن های پیش التهابی NOS2، I-6، COX2 و I-12 در BMDM های WT تیمار شده با فلورتین تحریک شده با LPS مشاهده شد (شکل 2B, C). در رابطه با نقش حیاتی Nrf2 در القای فنوتیپ کمتر التهابی، دادههای ما نشان میدهد که فلورتین ژنهای پاسخدهنده Nrf{13}}HO1 و NQO1 را در WT فعال میکند، اما نه Nrf2 KO BMDMهای تحریکشده با LPS (شکل 2D). علاوه بر این، تیمار فلورتین تولید ROS را در WT کاهش داد اما در Nrf2 KO BMDMs (شکل 2E) را کاهش داد. علاوه بر کنترل پاسخ های آنتی اکسیداتیو، Nrf2 برای سرکوب فنوتیپ التهابی ماکروفاژها گزارش شده است [12]. در حمایت از دومی، فلورتین قادر به کاهش بیان ژن های پیش التهابی NOS2، IL{25}}، COX2 نبود. ، و IL-12 در BMDM های فعال Nrf2- دارای کمبود (شکل 2F). در مجموع، این نتایج نشان می دهد که Nrf2 باعث تغییر فنوتیپ التهابی ماکروفاژها با واسطه فلورتین می شود.
فلورتین باعث فعال شدن AMPK می شود
فلورتین یک مهارکننده GLUT است که به خوبی تعریف شده است و مطالعات اخیر اهمیت جذب گلوکز با واسطه GLUT را برای فعال سازی ماکروفاژها برجسته می کند [26،32]. از این رو، ما تعیین کردیم که آیا فلورتین می تواند حسگر انرژی AMPK را فعال کند، که در سطوح کم انرژی / گلوکز فعال می شود. نتایج ما نشان می دهد که درمان با فلورتین منجر به فسفوریلاسیون و فعال شدن AMPK می شود (شکل 3A, B). علاوه بر این، افزودن بازدارنده AMPK BML{5}} تا حد زیادی از فعال شدن AMPK در BMDMهای تیمار شده با فلورتین جلوگیری می کند (شکل 3A, B). حتی بیشتر، یافتههای ما نشان میدهد که فعالسازی AMPK برای فلورتین برای سرکوب تولید ROS ضروری است، همانطور که با سطوح بالاتر ROS در BMDMهای تیمار شده با فلورتین و مهارکننده AMPK در مقایسه با BMDMهایی که با فلورتین به تنهایی درمان میشوند نشان داده شده است (شکل 3C). در مجموع، دادههای ما نشان میدهد که فعالسازی AMPK ناشی از فلورتین برای سوق دادن ماکروفاژها به سمت فنوتیپ کمالتهابی بسیار مهم است.
. The dotted line represents control cells stimulated with LPS. C ROS production in phloretin-treated or phloretin and AMPK inhibitor-treated BMDMs (n = 10). The dotted line represents control cells stimulated with PMA. Ctrl, control; phl, phloretin. Data are represented as mean ± s.e.m. *p < 0.05, **p < 0.01 and ***p < 0.001")
فلورتین اتوفاژی را به روشی وابسته به AMPK تحریک می کند
از آنجایی که فعالسازی AMPK با فعالسازی فرآیندهای کاتابولیک در پاسخ به محرومیت از مواد مغذی مرتبط است [33]، در ادامه بررسی کردیم که آیا فلورتین میتواند اتوفاژی را فعال کند یا خیر. اتوفاژی یک فرآیند کاتابولیک حفاظت شده است که بر اثر گرسنگی و سایر پاسخهای استرس ایجاد میشود، که باعث تخریب لیزوزومی محمولههای درون سلولی میشود که در وزیکولهایی که اتوفاگوزوم نامیده میشوند. تجزیه و تحلیل RNA-seq اتوفاژی را به عنوان یکی از فرآیندهای بیولوژیکی پایین دستی پیش بینی کرد که نشان دهنده افزایش فعالیت در BMDM های تیمار شده با فلورتین است (z-score: 0.779) (شکل 4A). علاوه بر این، تجزیه و تحلیل ایمونوسیتوشیمی BMDM های تیمار شده با فلورتین و بازدارنده اتوفاژی بافیلومایسین Al تجمع نشانگرهای اتوفاژی MAP1LC3/LC3 (پروتئین مرتبط با میکروتوبول 1 زنجیره سبک 3) و p62 را در مقایسه با بافیلومایسین A (شکل BMDMs) افزایش داد. . 4B-D)، نشان دهنده افزایش شار اتوفاژیک پس از درمان با فلورتین است [34]. با القای اتوفاژی، LC3 از فرم LC3I به فرم لیپید شده LC3I تبدیل می شود که با تعداد اتوفاگوزوم ها ارتباط دارد. در راستای این، سطوح پروتئین بالاتر LC3II و p62 در BMDM های تحریک شده با فلورتین توسط وسترن بلات تعیین شد (شکل 4E، F). جالب توجه است که این افزایش p62 و LC3II توسط فلورتین با درمان BMDM با مهارکننده AMPK BML{27}} (شکل 4E-F) جلوگیری شد. در مجموع، داده های ما نشان می دهد که فلورتین اتوفاژی را به شیوه ای وابسته به AMPK تحریک می کند.
. Data are represented by a heat map containing the normalized counts of genes associated with autophagy. A colour gradient was used to indicate the normalized counts and corresponding Fold change (Fc) differences, per sample and gene respectively. B–D Quantification and representative images of LC3 and p62 staining in phloretin-treated BMDMs stimulated with bafilomycin A1 (n = 5). The dotted line represents untreated cells (controls). E, F Western blot quantification and representative bands of the autophagy markers LC3II and p62 in phloretin-treated BMDMs stimulated with bafilomycin A1 alone or bafilomycin and the AMPK inhibitor together (n = 2). The dotted line represents cells treated only with bafilomycin A1 (controls). Ctrl, control; phl, phloretin; baf, bafilomycin A1. Data are represented as mean ± s.e.m. *p < 0.05 and **p < 0.01")
فلورتین مسیر Nrf2 را از طریق تخریب Keap1 با واسطه اتوفاژی فعال می کند
داده های ما نشان می دهد که فلورتین Nrf2 را فعال می کند و اتوفاژی را در ماکروفاژها تحریک می کند. جالب توجه است که گیرنده اتوفاژی p62 می تواند با Nrf2 برای اتصال به Keapl رقابت کند، آداپتوری که یوبی کوئیتیناسیون و تخریب Nrf2 را در شرایط عادی تسهیل می کند. بنابراین این p{4}}تفکیک میانجی Keap1/Nrf2 از تخریب Nrf2 جلوگیری میکند و در نهایت منجر به فعالسازی Nrf2 میشود [35]. به همین دلیل، ما تعیین کردیم که آیا فلورتین تعامل p62 / Keapl را ترویج می کند، در نتیجه مسیر Nrf2 را فعال می کند. در اینجا، نشان میدهیم که فلورتین مسیر Nrf2 را از طریق تخریب Keapl با واسطه p{13} در ماکروفاژها فعال میکند. با استفاده از میکروسکوپ کانونی Airyscan با وضوح بالا همراه با تجزیه و تحلیل colocalization، نشان میدهیم که فلورتین برهمکنش p62 و Keapl را تحریک میکند، همانطور که با افزایش مقدار پارامتر colocalization (ضریب Pearson) در BMDMهای تیمار شده با فلورتین نشان داده شده است (شکل 5DA). ، ب). علاوه بر این، یافتههای ما نشان میدهد که سطوح پروتئینی پایینتری از Keapl در BMDMهای تیمار شده با فلورتین وجود دارد، که تخریب Keapl را تایید میکند (شکل 5C). برای تأیید اینکه تخریب وابسته به اتوفاژی است، ما بازدارنده اتوفاژی بافیلومایسین A1 را به تیمار شده با فلورتین اضافه کردیم. BMDM ها جالب توجه است که این کاهش در Keapl با درمان بافیلومایسین Al معکوس شد (شکل 5C). این نتایج با وسترن بلات تایید شد، نشان دهنده کاهش سطح پروتئین Keapl در BMDM های تیمار شده با فلورتین است که توسط بافیلومایسین A1 جلوگیری شد (شکل 5D، E).
 on control or phloretin-treated BMDMs (90+ cells per well, 3 wells). C Quantification of Keap1 positive counts in phloretin-treated BMDMs treated with or without bafilomycin A1 (90+ cells per well, 3 wells). The dotted line represents corresponding control cells stimulated with or without bafilomycin A1. D, E Western blot quantification and representative bands of phloretin-treated BMDMs treated with or without bafilomycin A1 (n = 4). The dotted line represents the corresponding control cells stimulated with or without bafilomycin A1. Ctrl, control; phl, phloretin; baf, bafilomycin A1. Data are represented as mean ± s.e.m. *p < 0.05 and **p < 0.01")
Phloretin reduces neuroinflammation in the EAE model To validate our findings in vivo, we investigated the impact of phloretin on the EAE model, the most commonly used animal model of MS, that is characterized by a pronounced macrophage-mediated inflammatory response in the CNS 5]. EAE animals treated with phloretin before disease onset showed reduced clinical scores compared to vehicle-treated animals (Fig. 6A). Importantly, even in a therapeutic setup, in which phloretin treatment was started after disease onset (clinical score>1)، فلورتین شدت بیماری را کاهش داد (شکل 6B). کاهش شدت بیماری در حیوانات تحت درمان با فلورتین با کاهش بیان ژن های التهابی MHCI همراه بود.
CD{0}}، Nos2، TNFa، IL-6، CCL4، CCL5 و CXCL2 در نخاع (شکل 6C). علاوه بر این، مقدار کاهش یافته سلول های F4/80 در نخاع حیوانات تحت درمان با فلورتین یافت شد (شکل 6E، F). از آنجایی که هم F4/80*microglia و هم F4/80* ماکروفاژها به التهاب عصبی کمک می کنند، تجزیه و تحلیل عمیق تر نشان داد که فلورتین به طور قابل توجهی مقدار F480 به اضافه TMEMl را کاهش می دهد{17}}
ماکروفاژها در موش EAE (تکمیلی شکل 2 DF) در حالی که روند غیر قابل توجهی به سمت کاهش مجدد در F480 به علاوه TMEMl19 به علاوه میکروگلیا مشاهده شد. مطابق با این یافته، فلورتین تولید واسطه های التهابی در میکروگلیا را سرکوب کرد اما این سرکوب در مقایسه با ماکروفاژها کمتر مشخص بود (شکل 2 تکمیلی AC). این یافته ها نشان می دهد که بهبود EAE توسط فلورتین عمدتاً ماکروفاژ محور است. فلورتین علاوه بر کاهش بیان ژنهای التهابی، بیان عوامل ضد التهابی و نوروتروفیک مانند IL{10}}، CNTF و IGF{11}} را در نخاع حیوانات EAE افزایش داد (شکل 6D. ). این یافته ها قویاً نشان می دهد که فلورتین با هدایت ماکروفاژها به سمت فنوتیپ حل کننده بیماری، شدت بیماری EAE را کاهش می دهد. مطابق با یافتههای قبلی در شرایط آزمایشگاهی، ما همچنین افزایش سیگنال Nrf2 را در CNS حیوانات EAE تحت درمان با فلورتین شناسایی کردیم که با افزایش بیان mRNA Nrf2 و اهداف پاییندست NQO1 و GPX1 نشان داده شد (شکل 6G). در مجموع، این داده ها نشان می دهد که فلورتین التهاب عصبی را هم در یک محیط پیشگیری و هم در زمینه درمانی سرکوب می کند. علاوه بر این، یافته های ما نشان می دهد که فلورتین ممکن است با سرکوب ویژگی های التهابی ماکروفاژها از طریق فعال سازی Nrf2، التهاب عصبی را کاهش دهد.
 B Disease scores of EAE mice treated with vehicle or phloretin on a daily basis after disease onset (disease score > 1) (therapeutic setup, 50 mg/kg ip, n = 5). C, D Quantitative PCR was used to determine the mRNA levels of the pro-inflammatory genes MHCII, CD86, NOS2, TNFα, IL6, Ccl4, Ccl5 and CXCL2 and the anti-inflammatory and neurotrophic genes IL-4, CNTF and IGF-1 in the spinal cord of phloretin-treated EAE animals (prophylactic setting). E, F Quantification and representative images of F4/80 staining on spinal cord tissue obtained from EAE animals treated with vehicle or phloretin in the prophylactic setting. G Quantitative PCR was used to determine the mRNA levels of genes related to the Nrf2 pathway (Nrf2, NQO1, GPX1) in the spinal cord of phloretin-treated EAE animals (prophylactic setting). Gene expression was corrected for the number of F4/80+ cells. Ctrl, control; phl, phloretin. Data are represented as mean ± s.e.m. *p < 0.05 and **p < 0.01")
")
بحث
در این مطالعه، نشان میدهیم که فلورتین ماکروفاژها را به روشی وابسته به Nrf{1}}به سمت فنوتیپ کمالتهابی سوق میدهد. فعال سازی اتوفاژی وابسته به AMPK و تخریب متعاقب آن Keapl زمینه ساز فعال سازی Nrf2 توسط فلورتین است. علاوه بر این، ما اثرات ضد التهابی فلورتین را در یک مدل EAE تأیید می کنیم که در آن شدت بیماری را کاهش می دهد و وابسته به ماکروفاژ را کاهش می دهد.التهاب عصبی.
ما نشان می دهیم که فلورتین فنوتیپ التهابی ماکروفاژها را سرکوب می کند. این مطابق با یافته های Wei-Tien Chang و همکاران است. که نشان داد فلورتین دارای ویژگی های ضد التهابی در رده سلولی ماکروفاژ است [36]. ما نشان میدهیم که القای این تغییر فنوتیپ به Nrf2 وابسته است. Nrf2 یک تنظیم کننده اصلی پاسخ های آنتی اکسیدانی است و فعال سازی آن به عنوان سوق دهنده ماکروفاژها به سمت یک فنوتیپ ضد التهابی شناخته شده است [1l, 12, 37]. علاوه بر این، چندین مطالعه تعریف کردند که فعال سازی Nrf2 التهاب عصبی و تخریب عصبی را در اختلالات CNS کاهش می دهد [{10] }}]. با این حال، اگرچه یافتههای ما نشان میدهد که Nrf2 زیربنای تغییر فنوتیپ ماکروفاژهای تیمار شده با فلورتین است، دادههای توالییابی RNA نشان داد که در میان فعالسازی Nrf2، مسیرهای دیگر تنظیم مثبت شدند. به طور خاص، تنظیم مثبت مسیر PPAR به دلیل ویژگیهای ضد التهابی و گفتگوی متقابل با Nrf2 [{17}}] مورد توجه است. علاوه بر این، از آنجایی که فلورتین یک مهارکننده GLUT به خوبی تعریف شده است و تغییر به گلیکولیز یک رویداد متابولیکی مهم برای فعال شدن ماکروفاژها است، تغییرات فنوتیپ نیز ممکن است تا حدی توسط اثرات متابولیکی مستقیم فلورتین بر این ناقلان گلوکز ایجاد شود [46،47]. تحقیقات بیشتری برای تعریف ارتباط PPAR در فعال سازی Nrf2 ناشی از فلورتین و اینکه تا چه حد تعدیل ایمنی ناشی از فلورتین تحت تأثیر تغییرات مستقیم متابولیک است، ضروری است. به طور کلی، یافتههای ما به وضوح نشان میدهد که فعالسازی Nrf2 نقش اساسی در تغییر فنوتیپ با واسطه فلورتین ایفا میکند.
یافته های ما نشان می دهد که فعال سازی Nrf2 توسط فلورتین توسط فعال سازی AMPK واسطه می شود. AMPK نقش مهمی در بازگرداندن هموستاز انرژی سلولی در مورد تنشهایی دارد که ATP را کاهش میدهد و در نتیجه نسبت ADP: ATP را افزایش میدهد، در نتیجه مسیرهای کاتابولیک جایگزینی را که ATP تولید میکنند روشن میکند، در حالی که مسیرهای آنابولیک را که ATP مصرف میکنند خاموش میکند [48]. بر همین اساس، فلورتین یک مهارکننده GLUT به خوبی تثبیت شده است و سطح گلوکز سلولی را کاهش می دهد [49-51]. چندین مطالعه نشان داد که فعال شدن AMPK در ماکروفاژها باعث ایجاد یک فنوتیپ سرکوبگر سیستم ایمنی می شود که نتایج ما را تایید می کند که نشان می دهد فعال سازی AMPK برای فلورتین برای کاهش التهاب ضروری است [52،53]. دادههای ما همچنین با مطالعات قبلی مطابقت دارد که نشان داده شده است فلورتین AMPK را در سایر انواع سلولها از جمله سلولهای چربی و سلولهای اندوتلیال فعال میکند [50، 54-56]. فلورتین ممکن است به طور غیرمستقیم AMPK را با افزایش AMP و کاهش سطح ATP فعال کند، زیرا AMP و ATP به ترتیب فعالسازی AMPK را تحریک یا مهار میکنند [48، 57]. با این وجود، CaMKK که یک کیناز در بالادست AMPK است، میتواند فعالسازی AMPK را در پاسخ به افزایش سطوح سلولی Ca2 پلاس بدون هیچ تغییری در سطوح AMP/ATP ترویج کند [58]. علاوه بر این، با توجه به شبکه گسترده تعامل بین AMPK و مسیرهای بالا و پایین دست، تحقیقات بیشتری برای روشن کردن مکانیسم اساسی فعالسازی AMPK ناشی از فلورتین مورد نیاز است.
داده های ما نشان می دهد که فعال سازی AMPK با واسطه فلورتین باعث تحریک اتوفاژی در ماکروفاژها می شود. همانطور که در بالا ذکر شد، فعال سازی AMPK یک فرآیند خود محافظتی است که هدف آن بازگرداندن تعادل انرژی سلول است [48]. اگرچه هیچ مطالعه ای نشان نداده است که فلورتین قادر به تحریک فنوتیپ ماکروفاژها با ترویج اتوفاژی باشد، با این وجود برخی مطالعات اثر فلورتین را بر مسیرهای اتوفاژی در سلول های سرطانی و سلول های کبدی نشان داده اند [59-61]. علاوه بر این، مطالعات مختلف نشان میدهد که فرآیندهای کاتابولیک پاییندست فعالسازی AMPK میتوانند اتوفاژی را فعال کنند [60، 62،63]. جالب توجه است، در پاسخ به گرسنگی گلوکز، اتوفاژی با واسطه AMPK توسط فسفوریلاسیون آغازگر اتوفاژی وارداتی unc-51 مانند کیناز فعال کننده اتوفاژی [64،65] القا می شود. با توجه به این، ما حدس می زنیم که AMPK اتوفاژی را برای بازیابی ATP از اجزای سلولی در پاسخ به کاهش گلوکز ناشی از فلورتین تحریک می کند. با این حال، گرسنگی گلوکز همچنین میتواند اتوفاژی را به شیوهای مستقل از AMPK فعال کند [66-68]. بنابراین، تحقیقات بیشتری برای تعیین اینکه آیا فعال سازی اتوفاژی توسط فلورتین نیز تا حدی مستقل از AMPK رخ می دهد، ضروری است.
مطالعات اخیر اهمیت اتوفاژی را برای هدایت فنوتیپ ماکروفاژ عملکردی نشان داد. در این زمینه، اختلال در تنظیم اتوفاژی در ماکروفاژها به طور مشخص با شروع آترواسکلروز و بیماریهای عصبی مرتبط است [69-72]. در اینجا، پیوندی بین درمان فلورتین، القای اتوفاژی، و فعالسازی Nrf2 ارائه میکنیم و نشان میدهیم که فلورتین فعالسازی Nrf2 را از طریق p{4}}تخریب میانجی Keap1 ترویج میکند. تا همین اواخر، افزایش p62 puncta به عنوان نشانه ای از کاهش اتوفاژی در نظر گرفته می شد زیرا p62 در اتوفاگوزوم ها با فعال شدن اتوفاژی تخریب می شود [34]. در رابطه با این، تجمع p62 با اختلالات اتوفاژی در ضایعات آترواسکلروتیک مرتبط است [73]. با این حال، این تصور در سال های اخیر به چالش کشیده شده است و اکنون تصور می شود که p62 برای تشکیل اتوفاگوزوم ها ضروری است و افزایش سطح p62 به موازات افزایش LC3Il puncta می تواند نشانه ای از القای اتوفاژی باشد [74]. بنابراین، همراه با ظرفیت فلورتین برای فعال کردن AMPK و نقش شناخته شده آن در فعال سازی اتوفاژی، نتایج ما نشان می دهد که تجمع P62 و LC3 در درمان بافیلومایسین A1 در ماکروفاژهای تحریک شده با فلورتین به دلیل افزایش شار اتوفاژیک است نه به دلیل اختلال. خویشتن خواری. جالب توجه است که لی وای و همکاران. اخیراً نشان داده شده است که علاوه بر فعال کردن Nrf2، اتصال p62 به Keapl نیز در تشکیل اتوفاگوزوم نقش دارد، که نشان میدهد فعالسازی اتوفاژی با واسطه فلورتین مستقیماً با افزایش فعالیت p62 از یک طرف و همچنین با تعامل بیشتر Keapl-p62 از طرف دیگر مرتبط است. 75]. Keapl یک آداپتور یوبیکوئیتین لیگاز مبتنی بر کولین{27}} است که در شرایط هموستاتیک یک همودایمر با Nrf2 تشکیل میدهد و در نتیجه یوبیکوئیتیناسیون Nrf2 و تخریب پروتئازومی را تقویت میکند [76]. اختلال در کمپلکس Keapl-Nrf2 منجر به تثبیت Nrf2 و انتقال به هسته می شود [77، 78]. به خوبی مستند شده است که اختلال در این کمپلکس از طریق اصلاح باقیماندههای سیستئین Keapl توسط مولکولهای الکتروفیل، برهمکنش مستقیم مولکولهای کوچک با Keapl یا تخریب Keapl با واسطه p{36} رخ میدهد [35]. Ying Y و همکاران یک برهمکنش مستقیم فلورتین و Keapl را بر اساس آزمایشهای سیلیکونی پیشنهاد میکنند که سپس منجر به فعال شدن مسیر Nrf2 در رده سلولی قلب میشود [79]. در اینجا، ما یک مکانیسم اضافی مرتبط با اتوفاژی ایجاد کردیم که توسط آن فلورتین Nrf2 را فعال میکند.
با استفاده از مدل EAE، ما ثابت کردیم که فلورتین التهاب عصبی را در داخل بدن کاهش می دهد. داده های ما قویاً نشان می دهد که فلورتین EAE را با سرکوب پاسخ التهابی به واسطه ماکروفاژها و فعال کردن مسیر Nrf2 بهبود می بخشد. در سایر مدلهای بیماری، فلورتین نیز دارای ویژگیهای محافظ عصبی و تعدیلکننده ایمنی است. مشخص شد که فلورتین مسیر Nrf2 را در مغز بر اثر ایسکمی مغزی فعال می کند و تجمع بتا آمیلوئید را در مدل بیماری آلزایمر موش کاهش می دهد [80-84]. فلورتین همچنین ممکن است سایر سلول های ایمنی را در داخل بدن تحت تأثیر قرار دهد، زیرا مشخص شده است که این ترکیب فعال سازی سلول های T و دندریتیک را در شرایط آزمایشگاهی مهار می کند [85، 86]. با توجه به اینکه التهاب عصبی در مدل EAE صرفاً وابسته به ماکروفاژ نیست، جالب است که تعریف کنیم تا چه اندازه کاهش التهاب عصبی توسط سایر سلولهای ایمنی انجام میشود. با توجه به این، اگرچه دادههای ما نشان میدهد که بهبود EAE عمدتاً توسط ماکروفاژها هدایت میشود و کمتر به مدولاسیون میکروگلیا وابسته است، آزمایشهای بیشتری برای روشن شدن میزان تأثیر فلورتین بر میکروگلیا در بیماریهای التهابی عصبی مورد نیاز است. به طور کلی، یافته های ما نشان می دهد که فلورتین با تأثیر بر فنوتیپ ماکروفاژها، التهاب عصبی را کاهش می دهد و پتانسیل درمانی فلورتین را برای اختلالات التهابی عصبی نشان می دهد.

نتیجه گیری
مطالعه ما نشان می دهد که فلورتین یک عامل تعدیل کننده ایمنی قوی است که خواص التهابی ماکروفاژها را در اختلالات التهابی عصبی تعدیل می کند. فعالسازی مسیر Nrf2، با واسطه فعالسازی اتوفاژی وابسته به AMPK و تخریب بعدی Keapl، برای هدایت این تغییر فنوتیپ ایجاد شد. از این رو، فلورتین یک عامل طبیعی امیدوارکننده است که می تواند برای کاهش بار التهابی بیماری های عصبی التهابی مانند ام اس مورد استفاده قرار گیرد.
منابع
1. Zeng Y، Peng Y، Tang K، Wang YQ، Zhao ZY، Wei XY، و همکاران. دی هیدرومیریستین تشکیل سلول های کف را از طریق جریان کلسترول وابسته به LXRalpha-ABCA1/ABCG در ماکروفاژها بهبود می بخشد. Biomed Pharmacother. 2018؛ 101:543-52. https://doi.org/10.1016/j.biopha.2018.02.124.
2. Xia M، Hou M، Zhu H، Ma J، Tang Z، Wang Q، و همکاران. آنتوسیانین ها جریان کلسترول را از ماکروفاژهای صفاقی موش القا می کنند: نقش گیرنده فعال شده توسط پراکسی زوم {گاما} - گیرنده X کبد {آلفا} - مسیر ABCA1. جی بیول شیمی. 2005؛ 280 (44): 36792-801. https://doi.org/10.1 074/jbc.M505047200.
3. Chang YC، Lee TS، Chiang AN. کوئرستین بیان ABCA1 و جریان کلسترول را از طریق مسیر وابسته به ap در ماکروفاژها افزایش می دهد. J Lipid Res. 2012؛ 53 (9): 1840-50. https://doi.org/10.1194/jlr.M024471.
4. Grajchen E, Hendriks JJA, Bogie JFJ. فیزیولوژی فاگوسیت های کف آلود در مولتیپل اسکلروزیس Acta Neuropathol Commun. 2018؛ 6 (1): 124. https://doi. org/10.1186/s40478-018-0628-8.
5. بوگی جی اف، استینیسن پی، هندریکس جی جی. زیر مجموعه ماکروفاژها و میکروگلیا در مولتیپل اسکلروزیس اکتا نوروپاتول. 2014؛ 128 (2): 191-213. https://doi.org/1 0.1007/s00401-014-1310-2.
6. Baecher-Allan C، Kaskow BJ، Weiner HL. مولتیپل اسکلروزیس: مکانیسم ها و ایمونوتراپی نورون. 2018؛ 97 (4): 742-68. https://doi.org/10.1016/j. neuron.2018.01.021.
7. Bogie JFJ، Grajchen E، Wouters E، Corrales AG، Dierckx T، Vanherle S، و همکاران. استئاروئیل کوآ دساتوراز{2}} خواص ترمیمی ماکروفاژها و میکروگلیاها را در مغز مختل می کند. J Exp Med. 2020؛ 217 (5).
8. Bogie JF، Stinissen P، Hellings N، Hendriks JJ. ماکروفاژهای فاگوسیتوز کننده میلین، تکثیر سلول های T خود واکنشی را تعدیل می کنند. J التهاب عصبی. 2011؛ 8 (1): 85. https://doi.org/10.1186/1742-2094-8-85.
9. Bogie JF, Timmermans S, Huynh-Thu VA, Irrthum A, Smeets HJ, Gustafsson JA, et al. لیپیدهای مشتق شده از میلین فعالیت ماکروفاژها را با فعال شدن گیرنده X کبد تعدیل می کنند. PLoS One. 2012؛ 7 (9): e44998. https://doi.org/10.1371/ journal.pone.0044998.
10. Bogie JF, Jorissen W, Mailleux J, Nijland PG, Zelcer N, Vanmierlo T, et al. میلین با فعال کردن PPARها، فنوتیپ التهابی ماکروفاژها را تغییر میدهد. Acta Neuropathol Commun. 2013؛ 1 (1): 43. https://doi.org/10.1186/2 051-5960-1-43.





